العصر: الطباشيري المبكر–المتأخر, 119–99 مليون سنة | |
|---|---|
 هيكل عظمي منصوب معروض في اليابان
| |
| المرتبة التصنيفية | نوع |
| التصنيف العلمي | |
| المملكة: | الحيوانات |
| الشعبة: | الحبليات |
| الطائفة: | الصوروبسيدات |
| الطويئفة: | الديابسيدات |
| الرتبة العليا: | الديناصورات |
| الرتبة: | سحليات الورك |
| الرتيبة: | صوربوديات |
| الفصيلة العليا: | ريباتشصورديات |
| الفصيلة: | ريباتشصورنيات |
| الجنس: | نايجرصورص |
| Paul Sereno | |
| الاسم العلمي | |
| Nigersaurus taqueti Paul Sereno ، 1999 |
|
| تعديل مصدري - تعديل | |
نايجرصورص (معناه «زاحف النيجر») هو جنس من الديناصور الصوربودي الريباتشصوردي، عاش خلال العصور الطباشيرية الوسطى قبل 115 إلى 105 ملايين سنة مضت خلال الحقبة الألبية. اكتشفت أحفورته في تكوين الرهاز في مكان يسمى بقدوفاوا في النيجر. حفريات الديناصور وصفت لأول مرة في بحث علمي نشر عام 1976، لكن أطلق عليه الاسم في 1999 بعد اكتشاف ووصف البقايا كاملةً. هذا الجنس يتضمن نوعا واحدا فقط وهو نايجرصورص تاكيتي تيمناً بعالم الأحياء القديمة الفرنسي فيليبي تاكيت المكتشف الأول لبقايا النايجرصورص.
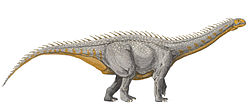
النايجرصورص طوله 9 أمتار (30 قدماً)، وهذا حجم صغير بالنسبة لصوربودي، وله رقبة قصيرة. تراوح وزنه حوالي الأربع أطنان، مماثل للفيل المعاصر. هيكله الهضمي متجوف بشدة (ممتلئ بالفراغات الهوائية المرتبطة بالحويصلات الهوائية)، لكن أطرافه ذات بنية وتركيبة قوية. جمجمته مخصصة بشكل كبير للتغذية مع ثقوب كبيرة وعظام رقيقة. له خطم عريض ممتلئ بأكثر من 500 سن كانت تتبدل بمعدل سريع حوالي كل 14 يوماً. ومن الممكن أن الفكين قد حملا غلافاً كيراتينياً. على النقيض من رباعيات الأطراف الأخرى، شكل فكاه ما يشبه المنقار العريض، وفكه أعرض من الجمجمة، والأسنان متموضعة بعيداَ عن مقدمة الجمجمة.
أقرب أقرباء النايجرصورص يصنفون ضمن فصيلة الريباتشصورنيات من فيلق الريباتشصورديات المتفرع من فصيلة الديبلودوكيديات الصوربودية. النايجرصورص كان على الأغلب باحث/مستكشف، وكان يتغذى ورأسه قريب من الأرض. المنطقة التي تكشف الروائح في دماغه كانت غير متطورة، ولكن على الرغم من هذا فحجم دماغه كان مماثلاً لأدمغة الديناصورات الأخرى. كان هناك جدال حول ما إن كان رأسه منخفض للأسفل بشكل فطري، أو أنه أفقي كالصوربودات الأخرى. سكن في موطن على ضفاف الأنهار، ونظامه الغذائي على الأرجح تألف من النباتات اللينة، كالسراخس، والكنباث، وكاسيات البذور. يعد واحدا من أكثر الأحفوريات الفقارية انتشارا في مناطق ضفاف الأنهار، وتشارك موطنه ديناصورات ضخمة أخرى، إضافة إلى الثيروبودوات والكروكوديلومورفيات الضخمة.
الأوصاف

كجميع الصوربوديات، النايجرصورص كان أحد رباعيات الحركة الضخمة صغيرة الرأس، مع سيقان خلفية غليظة، وذيل بارز. وسط هذا الفرع حيوي، النايجرصورص كان إلى حد ما صغير، مع جسد بطول 9 أمتار (30 قدماً) وعظم فخذ بطول يصل إلى 1 متر (3 أقدام). من الممكن أن وزنه كان يصل إلى 4 أطنان، مماثل إلى حد ما للفيل المعاصر. حظي برقبة قصيرة بالنسبة إلى صوربودي، مع ثلاثين فقرة عنقية. تقريباً جميع الريباتشصورديات حظيت برقبة قصيرة وطول يقدر ب10 أمتار (33 قدماً) أو أقل. الفرد الوحيد من عائلة الريباتشصورديات الذي حظي بحجم صوربوديات الأكبر كان هو الريباتشيصورص.[1]
الجمجمة
.jpg)
كان النايجرصورص يمتلك جمجمة رقيقة، مع ثقوب جمجمية (فتحات في الجمجمة) رباعية الأطراف أكبر من بقية الصوربوديات. كانت المساحة الكلية للعظم الرابط للخطم بمؤخرة الجمجمة تساوي 1.0 سـم2 (0.16 بوصة2). هذه الدعامات العظمية الرابطة تقدر في أغلب الأحيان بسمك أقل من 2 ميليمتر (0.08 إنش). بالرغم من ذلك، هذه الجمجمة كانت مقاومة للاقتصاص المتواصل للأسنان. وفيها أيضاً ميزة أخرى فريدة عن بقية الصوربوديات، إلا وهي امتلاكها ثقوب جمجمية مسدودة. الفتحات الأنفية، الخياشيم العظمية، كانت ممدودة.[1] وعلى الرغم من أن العظام الأنفية لم تكن معروفة تماماً، يظهر ان الحافة الأمامية للمنخر العظمي كانت أقرب للأنف من الديبلودوكيديات الأخرى. والأنف كان أيضاً أقصر بشكل متناسب، وصف الأسنان لم يكن فاقماً على الإطلاق، وطرف الأنف لم يكن ناتئاً مقارنة بباقي صف الأسنان.[2] صف الأسنان الفكي كان في استدارته العرضية الكاملة، جزئه الخلفي العادي مقلوب 90° تجاه المقدمة. وقابل هذا انقلاب مماثل له تماماً بمضرس الفك السفلي. وكنتيجة لم يمتلك أي رباعي أرجل كل أسنانه واقعة بكل هذا البعد عن مقدمة الجمجمة كالنايجرصورص.[1]

امتلكت الأسنان النحيلة قمم منحنية، والتي كانت إهليجية في المقطع العرضي. لربما كانت الأسنان في الفك السفلي أصغر ب20–30% من تلك التي في الفك العلوي، لكن القليل منها فقط معروف، والمعروف منها غير مؤكد نضجه. بمعزل عن هذا فالأسنان كانت بشكل عام متطابقة.[3] تحت كل سن نشط كان هناك طابور من تسعة أسنان بديلة ضمن الفك. مع 68 طابوراً في الفك العلوي و60 في الفك السلفي، هذه الملقبة بالبطاريات السنية (موجودة أيضاً في الهادروصوريات والسيراتوبسيا) تتألف كاملةً من أكثر من 500 سن نشط وبديل.[2] «البطاريات السنية» تنبت معاً بانسجام، ليس كل طابور ينمو بشكل منفرد.[3] المينا على أسنان النايجرصرص كانت «لا متماثلة» بشدة، كانت أكثر بعشر مرات أكثر على الجهة المقابلة للخارج أكثر من الجهة الداخلية من الفم.[4] هذه الحالة معروفة أيضاً في طيريات الورك المتقدمة.[3]
النايجرصورص لا يظهر نفس التغييرات على الفكين التي تحدث عند الديناصورات الأخرى مع البطاريات السنية، أو الثدييات مع وظائف مضغ مفصلية. كان الفك السفلي لديه يشبه شكل «حرف S اللاتيني» وانقسم على نفسه إلى رَّأْد مستعرض شبه إسطواني، والذي احتوى على الأسنان، والرأد الخلفي، وكان أكثر خفة وشكل الموقع لمعظم الأربطة العضلية. الفكين أيضاً أحتويا أعلى بضع ثقوب (نوافذ), من ضمنها ثلاثة فريدة من نوعها لا توجد في أي صوربودي آخر. النهاية الأمامية للفكين احتوت على الأخاديد التي تدل على وجود الغلاف الكيراتيني.[1] النايجرصورص هو الحيوان رباعي الأرجل الوحيد المعروف الذي امتلك فكين أعرض من الجمجمة وأسنان تمددت حرفياً على طول المقدمة. الأنف كان حتى أعرض من هؤلاء المنتمين إلى «بطيات المنقار» الهادروصوريات.[5]
تاريخ الاكتشاف

اكتشفت البقايا المَزْعَمُ انتماؤها إلى النايجرصورص لأول مرة خلال بعثة 1965-72 إلى النيجر تحت قيادة عالم الإحاثة الفرنسي فيليبي تاكيت، وذُكرت لأول مرة في ورقة نشرت عام1976.[2][6] على الرغم من أنه جنس شائع، إلا أن المعلومات المتوفرة عنه كانت قليلة جداً حتى تم اكتشاف عينات لأفراد آخرين من هذا الجنس خلال البعثة التي قادها عالم الإحاثة الأمريكي بول سيرينو من عام 1997 حتى عام 2000. الفهم المحدود لهذا الجنس نتج عن الحفظ الرديء للبقايا، والذي نتج عن الحالة الرقيقة والبنية الهوائية الشديدة (وجود عدد كبير من الثقوب الهوائية) للجمجمة والهيكل العظمي، ما سبب تفكك الحفريات. بعض أحافير الجماجم كانت رقيقة للغاية لدرجة أن شعاع الضوء كان مرئياً عبرها. وهكذا، لم يتم إيجاد أي جماجم سليمة أو هياكل عظمية بَيِّنة، وهذه العينات تمثل أكثر بقايا الريباتشصورديات المعروفة اكتمالاً.[1][2]

تمت تسمية ووصف النايجرصورص بشكل أكثر تفصيلاً من قبل سيرينو وزملائه في عام 1999, بناءً على بقايا الأفراد التي وجدت حديثاً. نفس المقالة ذكرت أيضاً جوباريا, ديناصور صوربودي آخر وجد في النيجر. اسم الجنس نايجرصورص («زاحف النيجر») هو إشارة إلى بلد اكتشاف المستحاثة، والاسم الدقيق تاكيتي سمي تكريماً لتاكيت، الذي كان أول من نظم بعثات إحاثية واسعة النظاق إلى النيجر.[3] عينة المزرعة النمطية (MNN GAD512) تتكون من جمجمة ورقبة جزئيتان. عينات لأطراف وعظام الكتف وجدت قريباً وعزيت أيضاً لنفس العينة. هذه الحفريات موجودة الآن في المتحف الوطني النيجيري.[2] سيرينو وجيفري ويلسون وفرا أول وصف مفصل للجمجمة والتكيفات الغذائية في 2005.[2] في 2007 تم نشر وصف أكثر تفصيلاً للهيكل العظمي، بناءً على عينة اكتشفت قبله بعشر سنوات. تم بعد ذلك عرض الحفريات بجانب نصب الهيكل العظمي المعاد بناؤه ونموذج بلاستيكي للجمجمة والرقبة في الجمعية الجغرافية الوطنية في واشنطن العاصمة.[7] لُقِّبَ النايجرصورص في الصحافة «ببقرة الحقبى الوسطى», وأكد سيرينو بأنه أكثر ديناصور إستثنائي رآه على الإطلاق. وقام بتشبيه مظهره الجسدي بدارث فيدر وبالمكنسة الكهربائية، وشبه قواطعه (الأسنان القاطعة) بالسير الناقل ومفاتيح البيانو المدببة.[7] تم إيجاد أسنان مشابهة لأسنان النايجرصورص في جزيرة وايت والبرازيل، ولكنه من غير المعرف ما إن كانت تنتمي لأقرباء لهذه الأصنوفة، أو انها تعود للتيتانوصور، الذي وجدت بقايا له في منطقة مجاورة. الفك السفلي لتايتانصور الأنتاركتوسورس يشبه إلى حد كبير الفك السفلي للنايجرصورص, لكنه من الممكن انه تطور تقاربياً.[2]
الصنف


تم وصف بقايا النايجرصورص في بادئ الأمر على أنها تنتمي إلى ديكراصورد، لكن في عام 1999 فريق سيرينو أعاد تصنيفه على انه أحد الريباتشصورديات الديبلودوكيدية.[3] الريباتشصورديات هي عائلة جذرية ضمن فصيلة الديبلودوكيديات، والتي تتضمن أيضاً الديبلودوكيد الطويل الرقبة والديكراصورد القصير الرقبة. فصيلة الريباتشصورنيات، التي تتضمن النايجرصورص وجنس آخر قريب، تمت تسميته من قبل جون وايتلوك عام 2011.[8] وفيما يلي مخطط تشعبي لتحليلات كتبها فيديريكو فانتي وزملاؤه عام 2013, والتي أكدت على أن النايجرصورص جذر رئيسي من فصيلة الريباتشصورنيات الريباتشصوردية.[9]
| الريباتشصورديات |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
تم وصف الجنس المقرب الديمانداصورص من إسبانيا في 2003, بجانب مجموعات حيوانية أخرى امتدت في إفريقيا وأوروبا العصر الطباشيري, هذا يشير إلى أن المنصات الكربونية ربطت هذه اليُبَّس عبر بحر البانثالاسا.[1] وتم دعم هذا الإدعاء في 2013 بوصف للتاتاوينيا الريباتشصورنية من تونس, والذي كان أقرب للأجناس الأوروبية أكثر من النايجرصورص, على الرغم من كونه أفريقياً, التي كانت جزءاً آنذاك من القارة العملاقة غندوانا.[9] قد يشير اكتشاف هذه الجذور الديبلودوكيدية أن الرقاب القصيرة والأحجام الصغيرة كانت مزايا سلفية للمجموعة.[1] حجيرات هيكل الريباتشيصورد العظمي الهوائية تطورت تدريجياً, وبلغت ذروتها في الريباتشصورنيات.[9] وفي دراسة تشعبية اجريت عام 2015, نسبت الريباتشيصورص إلى نفس مجموعة الريباتشصورنيات, النايجرصورص كان العضو الجذري في هذه الشعبة«الأفرو-أوروبية» الفرعية.[10]
علم الحفريات البيولوجية

تم إقتراح تصنيف النايجرصورص كديناصور أرضي, متصفح لاإنتقائي. عرض الخطم والتوجه الجانبي لصف الأسنان يظهر أن الصوربود يستطيع جمع الكثير من الطعام ويقوم بحصده على مسافة قريبة من الأرض, ضمن حدود متر واحد من السطح.[1][2] ويدعم هذا الإفتراض وجود احتكاكات على الجهة الشفوية (المواجهة للخارج) من الأسنان العلوية, في حالة مشابهة للديكراصورص والديبلودوكس, وهي دلائل على أن الطعام أَهْرَأَ أسنان الحيوان أثناء تغذيه. وعانى النايجرصورص أيضاً من علامات إهتراء سن إلى سن منخفض الزاوية في الجانب الداخلي من تيجان ثنايا الفك العلوي,وهذا يقترح أن حركة الفك كانت مقتصرة على الصعود والنزول. لم يتم اكتشاف أي اسنان سفلية (من الفك السفلي) مهترئة إلى الآن, ولكن من المتوقع إظهاره لإهتراء السن-للسن. المقدر على رفع رؤوسهم عالياً عن الأرض لا تعني بالضرورة أنهم بحثوا عن أية عناصر هناك, ورقبة النايجرصورص القصيرة قيدت مدى البحث بالمقارنة مع الديبلودوكيديات الأخرى.[1]

يبدو أن عضلة الفك المقربة كانت متصلة العضلة المربعة بدلاً من النافذة الفوق صدغية. كلاً منهما وباقي عضلات المضغ كانوا ضعافاً على الأرجح, ويُقدر أن النايجرصورص كان يملك قدرة مضغ من الأضعف بين أمثاله من الصوربوديات.[1] بالإضافة إلى تمتع الخدوش والحفر السنية بحجم صغير وطبيعة موازية تقريباً (والتي تسبب بها الحصى, والتي لا يتحصل عليها في العادة المتصفحات العالية) مما يشير إلى أنه أكل النباتا العشبية واللينة كالسرخس المنخفض النمو.[5] بسبب التوجه الجانبي للأسنان, على الأغلب انه لم يملك القدرة على المضغ إطلاقاً.[2] كانت أسنان النايجرصورص تهترئ أسرع من بقية الديناصورات الآكلة للأعشاب,[1] ومعدل تبدل الأسنان لديه كان الأسرع بين جميع الديناصورات المعروفة. كل سن كان يتم استبداله كل 14 يوماً; كان من المعتقد أن الصوربودات مع معدل تبدل أسنان أقل وتيجان أسنان أوسع كانت باحثات ظلة.[4]
على الرغم من امتلاكه فتحات انف كبيرة وخطم لحمي, إلا أن النايجرصورصامتلك منطقة شمية غير متطورة في الدماغ وهكذا فهو لم يمتلك حاسة شم متطورة. معدل كتلة الدماغ إلى الجسم عنده كان متوسطاً بالنسبة إلى سحلية, وأصغر من أولئك الثيروبودات اللا-كويلوروصورية. شكل مخه نسبة 30% من حجم الدماغ, كالكثير من الديناصورات الأخرى.[1]
وضعية الرأس

على أساس المسح المجهري لعناصر الجمجمة في عينة المزرعة النمطية, سيرينو وفريقه ابتكروا «نموذج» لجمجمة النايجرصورص لكي يستطيعوا إجراء الإختبارات عليها. هم أيضاً صنعوا قالب داخلي للدماغ وقاموا بمسح النفق هلالي في الأذن الداخلية ضوئياً, والتي وجدوها موجهة أفقياً. في دراستهم التي قاموا بها في 2007, هم اشاروا إلى أن هيكل القذال والفقرات العنقية قاما بتقييد حركات الصعود والهبوط الرقبة وإستدارة الجمجمة. وبناءً على تحليل النشاط الحيوي هذا, الفريق خلص إلى أن الرأس والخطم كانا بالعادة موجهان 67° إلى الأسفل وقريبين مستوى الأرض, كتأقلم للتصفح على مستوى الأرض. هذه ليس نفس الطريقة التي تم بها استعادة بقية الصوربودات, فالآخرين كانت رؤوسهم مرفوعة أكثر أفقيةً.[1]
في دراسة قام بها مايكل تايلور وماثيو ويدل ودارين نايش عام 2009 وافقوا أن النايجرصورص كان قادراً على التغذي ورأسه وعنقه في وضعية موجهة للأسفل وهو الاقتراح الذي قدمته دراسة أخرى في 2007, ولكن ما تم الاختلاف حوله هو ما إن كان هذه الوضعية الفطرية لهذا الحيوان. وأشارت الدراسة إلى ان وضعية الرأس والرقبة «المحايدة» للحيوانات المعاصرة لا تطابق بالضرورة وضعيتهم الفطرية. وذكرت أيضاً ان توجه القنوات الهلالية يختلف اختلافاً كبيراً عن تلك التي في الأنواع المعاصرة, ومن ثم فهي ليست جديرة جداً لمقارنة وتحديد وضعية الرأس.[11] وتم دعم هذا الإقتراح بدراسة أخرى عام 2013, اقترحت أن الأساليب التي تم اسخدامها من سيرينو وفريقه كانت غير دقيقة, وأن النايجرصورص كان يحمل رأسه بوضعية مشابهة لبقية الصوربودات.[12]
علم البيئة القديمة

عُرف النايجرورص من تكوين الرحاز ومجموةع تيغاما في منطقة تسمى غادوفوا, في النيجر. تلك المنطقة هي أحد أكثر مناطق التكوين التي تم اكتشاف فقاريات فيها. يتشكل تكوين الرحاز بشكل رئيسي من الحجار الرملية الغرينية الغائرة, والتي يحجب معظمها الكثبان الرملية.[3][13] الرواسب حَبّية خشنة إلى متوسطة, بدون وجود أي أفق حُبَيّبية ناعمة. النايجرورص عاش منذ 115 إلى 105 ملايين عام مضت في ما يسمى اليوم بالنيجر, في الفترتين الأبتية والألبية خلال العصر الطباشيرية الوسطى.[3] على الأرجح أنه عاش في موطن دَمرته السهول الفيضية الداخلية (منطقة ضفافية).[1]
بعد الإغوانودونتيا واللوردوصورص النايجرورص كان أكبر الحيوانات ضخمة عدداً.[1] آكلات الأعشاب الأخرى في نفس التكوين (تكوين الرحاز) من ضمنها الاورانوصورص والالرحازوصورص وتيتانوصور غير مسمى. عاشت أيضاً جنباً إلى جنب مع ثيروبودات أخرى كالكريبتوبس والسوكوميمس والإيوكارشاريا ونواصوريدا غير مسمى بعد. وكروكوديلومورفيات كالساركوسوكاس والاناتوسوكاس والأراريبيسوكاس والستولوكروسوكاس أيضاً عاشت هناك. وأيضاً تم إيجاد بقايا تيروصور وسلحفاة وسمكة, وقرش الهيبودونتيدا وذوات الصدفتين هناك.[13] العشب لم يتطور حتى العصور الطباشيرية المتأخرة, موجداً السراخس والكنباث وكاسيات البذور (التي تطورت في العصور الطباشيرية الوسطى) كطعام محتمل للنايجرورص. من غير المحتمل ان الديناصور تغذى على المخروطيات والسيكاديات والنباتات مائية بسبب ارتفاعه وهيكله الصلب والقاسي وافتقاد الموطن الملائم على التوالي.[1]
انظر أيضًا
مراجع
ref name="NPR">Joyce, C. (25 نوفمبر 2013). "'Mesozoic Cow' Rises from the Sahara Desert". NPR. مؤرشف من الأصل في 2018-06-26. اطلع عليه بتاريخ 2007-11-16.</ref>
[5] Because of the lateral orientation of the teeth, it probably would not have been able to chew.[2] Nigersaurus wore its tooth crowns down faster than other dinosaurian herbivores,[1] and its tooth replacement rate was the highest of any known dinosaur. Each tooth was replaced once every 14 days; the rate had previously been estimated lower. In contrast to Nigersaurus, sauropods with lower tooth replacement rates and broader tooth crowns are thought to have been ظلة browsers.[4]
- ^ ا ب ج د ه و ز ح ط ي يا يب يج يد يه يو يز سيرينو، باول؛ ويلسون، جيفيري؛ ويتمر، لورنس؛ ويتلوك، جون؛ ماغا، عبداللاييه؛ آيدي، عمرو؛ روي، تيموثي (2007). "الهيكلية المفرطة في ديناصور من العصر الطباشيري". PLoS ONE. ج. 2 ع. 11: e1230. DOI:10.1371/journal.pone.0001230. PMC:2077925. PMID:18030355.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link). - ^ ا ب ج د ه و ز ح ط ي Wilson, J. A.; Sereno, P. C. (2005). "Structure and evolution of a sauropod tooth battery". في كريستينا كاري روجرز, and جيفري ويلسون (المحرر). The Sauropods: Evolution and Paleobiology (PDF). University of California Press. ص. 157–177. ISBN:0-520-24623-3. مؤرشف من الأصل (PDF) في 2016-03-03.
{{استشهاد بكتاب}}: صيانة الاستشهاد: أسماء متعددة: قائمة المؤلفين (link) - ^ ا ب ج د ه و ز Sereno، P. C.؛ Beck، A. L.؛ Dutheil، D. B.؛ Larsson، H. C.؛ Lyon، G. H.؛ Moussa، B.؛ Sadleir، R. W.؛ Sidor، C. A.؛ Varricchio، D. J.؛ Wilson، G. P.؛ Wilson، J. A. (1999). "Cretaceous sauropods from the Sahara and the uneven rate of skeletal evolution among dinosaurs". Science. ج. 286 ع. 5443: 1342–1347. DOI:10.1126/science.286.5443.1342. PMID:10558986.
- ^ ا ب ج D’Emic، M. D.؛ Whitlock، J. A.؛ Smith، K. M.؛ Fisher، D. C.؛ Wilson، J. A. (2013). Evans، A. R. (المحرر). "Evolution of high tooth replacement rates in sauropod dinosaurs". PLoS ONE. ج. 8 ع. 7: e69235. DOI:10.1371/journal.pone.0069235. PMC:3714237. PMID:23874921.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - ^ ا ب ج Whitlock، J. A. (2011). Farke، A. A. (المحرر). "Inferences of diplodocoid (Sauropoda: Dinosauria) feeding behavior from snout shape and microwear snalyses". PLoS ONE. ج. 6 ع. 4: e18304. DOI:10.1371/journal.pone.0018304. PMC:3071828. PMID:21494685.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - ^ Taquet, P. (1976). "Géologie et paléontologie du gisement de Gadoufaoua. (Aptien du Niger)" (PDF). Cahiers de paléontologie (بالفرنسية). Paris: 53. ISBN:2-222-02018-2. Archived from the original (PDF) on 2017-03-29.
- ^ ا ب Joyce, C. (25 نوفمبر 2013). "'Mesozoic Cow' Rises from the Sahara Desert". NPR. مؤرشف من الأصل في 2018-06-26. اطلع عليه بتاريخ 2007-11-16.
- ^ Whitlock، J. A. (2011). "A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda)". Zoological Journal of the Linnean Society. ج. 161 ع. 4: 872–915. DOI:10.1111/j.1096-3642.2010.00665.x.
- ^ ا ب ج Fanti، F.؛ Cau، A.؛ Hassine، M.؛ Contessi، M. (2013). "A new sauropod dinosaur from the Early Cretaceous of Tunisia with extreme avian-like pneumatization". Nature Communications. ج. 4. DOI:10.1038/ncomms3080.
- ^ Fanti، F.؛ Cau، A.؛ Cantelli، L.؛ Hassine، M.؛ Auditore، M. (2015). "New Information on Tataouinea hannibalis from the Early Cretaceous of Tunisia and Implications for the Tempo and Mode of Rebbachisaurid Sauropod Evolution". PLoS ONE. ج. 10 ع. 4: e0123475. DOI:10.1371/journal.pone.0123475.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - ^ Taylor، M. P.؛ Wedel، M. J.؛ Naish، D. (2009). "Head and neck posture in sauropod dinosaurs inferred from extant animals". Acta Palaeontologica Polonica. ج. 54 ع. 2: 213–220. DOI:10.4202/app.2009.0007.
- ^ Marugán-Lobón، J. S.؛ Chiappe، L. M.؛ Farke، A. A. (2013). "The variability of inner ear orientation in saurischian dinosaurs: Testing the use of semicircular canals as a reference system for comparative anatomy". PeerJ. ج. 1: e124. DOI:10.7717/peerj.124. PMC:3740149. PMID:23940837.
{{استشهاد بدورية محكمة}}: صيانة الاستشهاد: دوي مجاني غير معلم (link) - ^ ا ب Sereno، P. C.؛ Brusatte، S. L. (2008). "Basal abelisaurid and carcharodontosaurid theropods from the Lower Cretaceous Elrhaz Formation of Niger". Acta Palaeontologica Polonica. ج. 53 ع. 1: 15–46. DOI:10.4202/app.2008.0102.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||